
http://www.flash-screen.com/free-wallpaper/free,wallpaper,3426,html
The Dominance of the Dinosaur
Sauropods all, wherever you may be,
If you want to dine at the top of the tree
Where nectar’d flowers hang abundantly,
Keep your necks up high and never bend a knee
And you all may feast on Proteaceae.
(Apologies to W. S. G.)
Dr. Harry Levin finds the rhythms and rhymes of humor, even in prehistory. This is essay number five of 10 essays devoted to a basic re-examination of the late Paleozoic and Mesozoic eras of earth history, published for the first time. He describes the heyday of the dinosaur.
 Dr. Harry Levin |
Did
the giant Brachisaurus feast on the flowers of Proteaceae? Did
dinosaurs migrate in the same direction as birds of today? Dr. Levin, a chemical engineer, has contributed groundbreaking essays ranging from the Origin of Flowers to a new theory on how New Zealand is related to South America in deep time. He has presented a pictorial essay The Evolution of Proteaceae, in Flower and Leaf, and photographs of flowers that have attracted visitors from around the world. He has written about The Geological Significance of a Fish. Dr. Levin, of Woodland Hills, California, describes how his analytical process crosses the disciplines: |
The interweaving here of geophysics and biophysics – both embracing climatology – brings out clarifying and sometimes incontrovertible evidence – content otherwise obscure to, or hidden from, either one or the other of these disciplines when it acts alone."
Florida Wildflowers takes pride in presenting this essay.
– Michael E. Abrams
By Harry Levin
Copyrighted
Abstract
It is of the utmost significance to recognize that the dinosaur could float and swim in herds, as was its habit. Furthermore, the ascendancy of the dinosaur was closely associated with the presence of the angiosperm as food source, from the first dinosaurs to the earliest birds.
The instinctive migratory behavior of the dinosaur in search of safe nesting sites and near-by food was probably the key to its 160-million-year dominance. This essential migratory land-and-water behavior is presented here in context with the relevant paleogeography. Critical to its long survival was the ability that this specific pattern of migration provided the dinosaur to safeguard its eggs and young from predators.
Dinosaur survival in large measure was due to a fortunate geographical circumstance that gave rise to its seasonal migratory habit. Propinquity of Gondwanaland and Laurasia continental margins during the entire Mesozoic – their separation defined by a narrow oceanic channel termed here the “Western Tethys”– contributed remarkably to the 160-million-year dominance of the dinosaur upon the earth. This fortuitous geography – and the aquatic ability of the dinosaur – enabled the periodic crossing of the extensive narrow western reaches of a once vast expanse of Tethys Ocean.
North in season across the Western Tethys (Figure 1) went herds of dinosaurs into Laurasia to breed in large numbers, in safety from predators such as therapsids and early mammals. Then south again in new season went the herds into Gondwanaland to feed on plenteous angiosperms. Inherent cyclic north and south migratory patterns are characteristic of most birds today.
1
Figure 1
The
Western Tethys sea channel, Jurassic, 200 million years ago
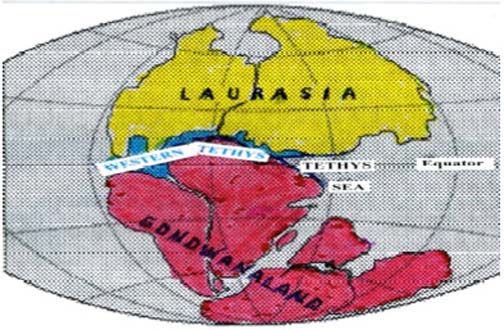
The
waterway is positioned in blue upon a U.S. Geographical Survey
chart authored by W. J. Klous and R.I. Tilling (1)
chart authored by W. J. Klous and R.I. Tilling (1)
Introduction
In hypothesis, dinosaur survival in large measure was due to favorable geographical circumstances, to wit: the separation of Gondwanaland and Laurasia continental margins during the entire Mesozoic by a narrow oceanic channel termed here the “Western Tethys.” (See the Figure 1)
For many reasons, the dinosaurs could cross this seaway; but therapsids (mammal-like reptiles) and mammals could not. The unique crossability of this channel by the dinosaurs contributed remarkably to the 160-million-year dominance of the dinosaur upon the earth. Of special significance to the dinosaur crossings was the ancient location of the angiosperm. The angiosperm was endemic solely on Gondwanaland soil until the late Jurassic, 150 million years ago.
Migratory herd behavior governed the existence of the dinosaur. This behavior abides today within the descendant bird. (The herd was father to the flock.) Herds of herbivore dinosaurs, attended by carnivore dinosaurs, the theropods, swam north in season across the Western Tethys to Laurasia to breed in safety away from therapsids and mammal predators. Then back again they went in season to Gondwanaland to feed on indigenous angiosperms. As a result of this ploy of safe breeding, the dinosaurs dominated by sheer number. Their theropods overwhelmed the therapsids and sent the mammals into hiding.
2
The Permian ended 245 million years ago with extinction of 70% or more of land creatures, by various estimates. Ecological opportunity then opened for a new reptile subclass, the dinosaur, to appear on the land during the early Triassic.
The epic demarche of dinosaurs into biogeophysical history was preceded by the therapsid and small-sized mammals. Long before the coming of dinosaurs, they lived in the cold temperate, angiosperm woodlands of Gondwanaland. Among them were wily predators that over millions of years of coexistence with other reptiles had acquired taste and skills for eating reptile eggs and young reptiles. Therapsids threatened the existence of the dinosaur from the first early Triassic encounters. (Placental mammals were a threat to dinosaur eggs and juveniles even during the Jurassic; but they became a severe honing force to dinosaur breeding only in the late Cretaceous.)
The oldest herbivore dinosaur fossils are of a prosauropod. Fossils of this archetypal dinosaur, uncovered in Madagascar, have been determined to be about 230 million years old. The site of the discovery is presumptive of the Gondwanaland origin of the dinosaurs. Fossil evidence indicates that the prosauropods were plant eaters, probably feeding from a bipedal stance. Their forelimbs were shorter than their hind limbs, and some had long, moderately flexible necks. They ranged in size from 2 to 7 meters. From mid-Triassic through the early Jurassic (230 to 180 million years ago), they were found in South America and Africa, and, highly significantly, in the northern hemisphere too, in North America, Europe, and even China – on both sides of the Western Tethys seaway.
Dinosaur dominance started with the prosauropod. It was due, in effect, to natural selection. The early prosauropods nested probably at random on both sides of the Western Tethys. On the north shore (across from Gondwanaland) they experienced safe nesting. They adapted to the north shore for breeding. By crossing to the north shore, they were able to protect clutches of fragile eggs and succulent juveniles from therapsids and mammals that could not cross the water.
During the late Triassic (about 215 million year ago), the Sauropoda evolved, probably descendents of the prosauropods and following them in migration and breeding habits. Both belonged to the suborder Sauropodomorpha. The sauropod looms large in many ways in the total account of the dinosaur. Great diversity and abundance came during the late Jurassic and early Cretaceous, about 160 to 100 million years ago. They were quadrupeds, plant eaters, notable for their enormous size (ranging to more than 40 m) and for their long flexible necks, small heads, and enormous tapering tails.
3
Ubiquity of the Sauropodomorpha and the Theropoda
As in the earlier case of the prosauropods, the same families of sauropods were present on both sides of the seaway of the Western Tethys, according to the fossil records of the Jurassic. They were present in North America and Europe, as well as across the water in Africa and South America. Here are indications of natant abilities – that both infra orders, the prosauropods and the sauropods, were capable of floating and swimming; and that, from the first, the dinosaur was designed by nature to be an aquatic terrestrial reptile. The sauropod carved an amazing ecological niche for itself. Here was a creature that in some species weighed more than 60 metric tons, had a brain the size of a rabbit’s, and yet managed to exist for 160 million years to the end of the Cretaceous.
At this point, it would be remiss not to introduce a principal player among the dinosaurs, the suborder Theropoda. In the middle Triassic appeared the earliest infra order of theropods, the herrerasaurs, perhaps slightly later than the prosauropods. From the start the theropods were bipedal (and remained bipedal even into birdhood.) Some were scavengers and some perhaps even omnivores, but by and large, they were killers, “bloody in tooth and claw.”
They culled the herds of herbivores. Wherever went the Sauropodomorpha, there went the Theropoda at the fringes of the herd; and nature presumably adjusted the numbers to keep a constant balance of herbivore to carnivore. The theropods, of hollow bone, were as capable at swimming as the sauropodomorphs (2).
4
The geological prologue to the dinosaurs began about 380 million years ago during the late Devonian with collision of the continental margins of North America and Gondwanaland (3). Gondwanaland had thrust northward, giving rise to the Ouachita Mountains (of Arkansas and Oklahoma) where the northern margin of South American Gondwanaland entered into collision with the south-facing margin of North America.
The lifting of the Appalachian branch of the Allegheny orogeny then followed; and, farther east, arose the Hercynide southern European mountains. In time, the northwest margin of Africa confronted the southeast-facing margin of North America and Europe. The Western Tethys, the aforementioned relatively stable, narrow transcontinental seaway, evolved between them by marginal contact.
These were the orogenic events that extended into the Permian, as continents in approach gave form to the Western Tethys. Here the door opened for the dinosaur, from its modest Triassic entry, for its long stay upon the earth.
Genesis of the Transcontinental Seaway, the Western Tethys
This thesis tells of the significance of the transcontinental seaway, the Western Tethys. It became an entity prior to the Mesozoic and played a role essential to that heyday of the dinosaur. After about a hundred million years of its existence, a major extension of the Western Tethys occurred. In the Cretaceous, the Arabian Plate approached Laurasia from Africa; and as a result of the near contact, the Western Tethys was extended to its maximum length of close continental approach.
It was then bound by North America, Europe, and the Middle East to the north and by South America, Africa, and Arabia to the south. Thus, a long, narrow seaway of varying width had been sculpted. It persisted through about 160 million Mesozoic years where in the deeper past had been a paleo-Tethys ocean.
The stability of the seaway had many contributing causes. Drifting of the “nomad” North America Plate (4) had been slowed by its Laurussian coalescence with Europe during the early Devonian. In the late Permian, this union became the vast geomass Laurasia by the joining of Siberia and Khazakstan to the northeast. It would experience other complicating impulses during the Mesozoic in adjusting to the relentless pressure of an intervening mid-Atlantic-ridge sea floor spreading, pushing it westward along its entire length in collision with the eastward-moving Pacific Plate.
In Gondwanaland too, Africa and South America were bound together until about 150 million years ago. South America itself, beyond its Fuegian margin, was restrained by Antarctica until 15 to 30 million years ago. The result was that during the Mesozoic, North America and Europe were in close relationship. South America and Africa began gradually to drift apart in the late Jurassic.
5
Throughout the entire Mesozoic, the Western Tethys played a sustaining role for the dinosaur. The Western Tethys spanned the entire 165 million years of the Mesozoic. As mentioned above, the channel in its later years lengthened considerably. In effect, rotation of the Arabian Plate to the margin of Laurasia, had extended the Western Tethys by about one-third. The Persian Gulf opened eastward into the waters of the Indian Ocean, and in actuality, the lengthened Western Tethys connected the Mediterranean to the Indian Ocean. The Western Tethys remained open all through the Mesozoic. An ichthyosaur could swim from one end to the other.
An excellent indicator of the true nature of the Western Tethys as a seaway are the vast petroleum reservoirs – roughly half the world supply – that were formed in places which are now on the Gulf Coast of the United States, in Mexico, in Venezuela, in Africa, in southeastern Europe, but mostly in deep basins in the western area of the Persian Gulf, both to the north and to the south. Oil and natural gas was formed under optimum conditions during the middle Cretaceous, as the deep seaway narrowed between the Eurasian Plate (fronted by small Iranian and Anatolian Plates and an approaching Arabian Plate), while receiving fine sediment off the land.
These petroleum reservoirs suggest that at the times of oil formation, the water circulation of the Western Tethys was poor and often oxygen deficient, especially during the middle Cretaceous. There must have been frequent episodes of stagnation, of plankton bloom, of lethal toxin release, of anoxia, and of intermittent, overwhelming die-offs of the entire marine ecosystem. Indeed, the Western Tethys was mostly a gutter of organic material and marine sediment reaching one-third the way around the world during the late Jurassic and the Cretaceous. The gutter washed down to the deeper end, to deep basins at the Persian Gulf. There, the favorable combination of geology and organic chemistry built a large share of the world’s oil and natural gas reserves.
Between die-offs, these waters were host to seaweed and plankton organisms including dinoflagellates and innumerable nektonic life forms – more lush by far than a Sargasso Sea. The waters hosted fish, crustaceans, and mollusks (notably ammonites and belemnites, abundant cephalopods, which became extinct about the same time as the dinosaur).
The existence of the Western Tethys was ended by land closure during the Oligocene (37 to 24 million years ago). It was then, with land uplift and sea recession, that the Arabian Plate was able to pinch off the waters of the Tethys entirely, thus sculpting the eastern banks of the Mediterranean Sea.
6
During the entire existence of the Western Tethys, both as seaway and embayment, that body of water presented a barrier to angiosperms and other Gondwanaland flora against crossing to the north. Of special significance for the dinosaur was the location of the angiosperm: solely on Gondwanaland soil until 150 million years ago. Meaningful to Mesozoic history was this fact: angiosperms did not cross the Tethys. To quote Venkata Rao, 1971(5): “. . . the transglobal Tethys sea . . . would prove a barrier against plant migration.” Heeding Rao, different intervening edaphic and climate conditions – and salt water – could have barred the angiosperms from Western Tethys approach and crossing into the northern hemisphere.
Likewise, therapsids and mammals were not able to cross (with the highly significant exception that tiny, scansorial mammals were able to cross in unique cooperation with dinosaurs, as told in a later essay). Waiting to welcome therapsids and mammals with open jaws were aquatic reptiles (along with huge cuttlefish and other large sea creatures). These reptiles effectively guarded the seashore and inland dinosaur nesting sites. These were the plesiosaurs, ichthyosaurs, and later mosasaurs, and probably certain among the dinosaurs themselves, the huge Spinosaurus and the crocodile-like Suchomimus. That stewardship ended suddenly 66 million years ago with the complete extinction of plesiosaurs, mosasaurs and dinosaurs.
Perhaps it was because of a need for protection against these formidable aquatic creatures that the dinosaurs early adapted to highly sophisticated herding. The young herbivores were at the center of the herd; the theropods were at the fringes, the largest and fiercest of them guarding the periphery. Thus they were able to cross the Western Tethys back and forth in season with relative impunity.
Dinosaurs in herds could cross the Western Tethys. The crossing back and forth of the Western Tethys by dinosaurs was the critical cause of dominance of dinosaurs upon the earth. Dinosaurs could cross; therapsids could not. Thus, a safe haven for uninhibited dinosaur population growth was established. In consequence, dinosaurs (with characteristically large litters) overwhelmingly became sole rulers of the land. Safe haven for dinosaur breeding, as provided by the Western Tethys, was a major factor in the complete extinction of the dinosaurs’ fiercest competitors: the large crocodile-antecedent crorotarsans by about 208 Ma and the therapsids, by about 145 Ma.
The Dinosaur, Both Terrestrial and Aquatic
It is essential to understand how the dinosaur, in herds, could, in sequence: leave Gondwanaland; float and swim the Western Tethys; nest on dry northern shores; return and feed once again on the lush vegetation of Gondwanaland. As an exemplar, consider the sauropods. They were capable long-distance swimmers, even among seaweeds. They were at ease in the generally calm waters, abetted by low body density and protected by size and by herding instinct.
Their long necks served a double purpose. In good season, in Gondwanaland, these necks would reach high up into tall angiosperm trees for soft, juicy flowers. In another season, the necks would facilitate travel over water. High out of the water and with a small head, they were like the prows of Viking ships.
7
Their necks could reach down and place the head under water perhaps to feed on underwater vegetation. The heads bore nares that were situated to advantage near the top of the skull and were well-suited to take in air while the sauropod skimmed and fed at the water surface. The long tapered tails also, perhaps partially out of the water, could serve both to steer and to defend with a mighty force and a whip-like crack. And aided by buoyancy, the legs – even with their huge thighbones – could serve as oars.
As a vehicle both for land and sea travel, the sauropod body itself appears to have been designed for weight economy and buoyancy, ashore and afloat. The backbone was well engineered for weight sparing and weight bearing.
Each vertebra had a large centrum cavity, and at the same time, had complex muscle and ligament attachments to compensate for loss in bone strength.
Wrote Carrol L. Fenton and Mildred A. Fenton about 1989 (6): “As sauropods became giants, therefore, they developed ‘excavated’ vertebrae, which seemed to be made up largely of holes. Actually, evolutionary changes omitted bone where it did no good but added it in bars and ridges that met every normal stress. The result was a structure much like that of a modern steel bridge, held together by ligaments and interlocking joints instead of by welds, rivets, or bolts.” The legs were adapted to support the animal’s weight on dry land. The ponderous, muscular body could walk without difficulty to feed in an angiosperm forest. And in due season, the huge boat of a body, possessed of an enormous gut, could be loaded with buoyant fat and inflated with air for even a many-day voyage.
The survival strategy of the dinosaur was brilliant, demonstrating the patient magic of evolution. It was a strategy undone in time by an earth surface in change and by an asteroid incursion. Its reconstruction as earth history pleads for the telling. The mammal remained a shadowy, small, but cunning player until the playing field tilted in its favor during the late Cretaceous.
Herds, Then Flocks, Crossed the Waters, Back and Forth
The migration cycle continues today among birds. As they have done in the past, numerous species of birds obey their genetic dinosaur breeding instincts and migrate in season north to nest. The flights are essentially due north, with some veering to allow for resting and feeding. By way of examples, intertidal areas and near-coastal wetlands of the Yellow Sea (shared by China, North Korea, and South Korea) support at least two million shorebirds that in season use the region during northward migration. Among these shorebirds, both the Mongolian and greater sand plovers spend summers nesting in central Asia; and then they migrate south to Africa and to Australia. Then there is Louisiana’s Atchafalaya Swamp. It is the northern nesting habitat of more than 170 southern bird species.
8
Envoi
The dinosaur is no more.
In a word, it became a bird.
Summary
Reproductive advantage, once established, can rank as the most powerful stratagem for ascendancy of a phylum, a class, an order, a family, a genus, and a species. The dinosaur is a prime example. Dinosaurs originated in Gondwanaland during the early Mesozoic. They dominated the earth throughout the Mesozoic by a successful stratagem of cyclic migration. To wit, in season, they crossed the Western Tethys north into Laurasia to nest safely (from egg predation); and, in cycle, they returned south to feed in Gondwanaland.
Hence, dinosaurs chanced upon a powerful reproductive advantage. In the Triassic, the safe-bred theropod carnivores, by the newfound sheer number of their own offence force, were able to decimate populations not only of large crocodile-like crorotarsans but also of the therapsids -- and eventually bring about the extinctions of both antagonists. The singular ability of the dinosaur to traverse waters shore to shore over increasingly long distances (even beyond the Western Tethys) established the dinosaur during the Mesozoic as the dominant animal upon the earth.
9
1. U.S. Geological Survey, GPO Stock No. 024-001-03601-1, W.J. Kious and R.I. Tilling,
This Dynamic Earth: The Story of Plate Tectonics, illustrations by J. Russell, Washington,
D.C., 2001.
2. Ezquerra, R., Doublet, S., Costeur, L., Galton, P. M., and Perez-Lorente,
F., Were non-avian theropod dinosaurs able to swim? Geology, 35 (6)
507-510 (2007) Geological Society of America.
3. Dalziel, I. W. D., Scientific American 272, 56-63, (1995).
4. Tarling, D. H. and Tarling, M. P., Continental Drift, A Study of the Earth’s Moving
Surface, G. Bell and Sons, London, 1971.
5. Rao, C. V., Botanical Monograph No. 6, Proteaceae, Council of Scientific and
Industrial Research, New Delhi, 1971.
6. Felton, C. L., and Felton, M. A., The Fossil Book, pp. 448-449 and 454, Doubleday,
New York, 1989; revised and expanded by others.