New
Zealand: a
Late-Paleozoic Part of South America
By Harry Levin
copyrighted
Abstract
In what may be termed “one of the grand puzzles of the earth’s
prehistory,” New Zealand and Chile, as a common native habitat, share
the earth’s oldest, most primitive flowering plants, side by side with
the earth’s most ancient conifers. New Zealand and Chile are now some
9000 km. apart.
In The Origin of Species,
in 1859, Charles Darwin wrote:
“New Zealand in its endemic plants is much more closely related to
Australia, the nearest mainland, that to any other region: and this is
what might have been expected; but it is also plainly related to South
America, which, although the next nearest continent, is so enormously
remote, that the fact becomes an anomaly.”
This grand anomaly is resolved by evidence presented herein, which
indicates that New Zealand was once
part of South America on a fully consolidated Gondwanaland during the
late Paleozoic more than 300 million years ago. A New Zealand block was
then adjoined to the southwestern coast of Chile. This essay
addresses the geological and biological relationships of New Zealand to
South America and to Australia from the late Paleozoic to the present.
1
|
Molecular Genetic Support of Devonian
Angiosperm Origin
The basic hypothesis of Devonian origin of angiosperms (before 360 mya)
is supported by studies of DNA These studies evince an early divergence
of angiosperms from a Devonian plant (seed fern). Brief statements
epitomizing five such studies:
• V. Troitsky et al. (1) indicate (in 1991) by rRNA sequence
comparisons
that both gymnosperms and angiosperms are monophyletic groups. The
genealogical splitting of gymnosperm and angiosperm lineages occurred
at least 360 million years ago. Magnoliales were the earliest
angiosperms. They are dicotyledons.
• W. F. Martin et al. (2) indicate (in 1993), by chloroplast and
nuclear
sequence data taken together, that angiosperms and gymnosperms were
separate lineages about 330 million years ago and that the separation
of monocotyledonous and dicotyledonous lineages of angiosperms took
place about 300 million years ago.
• T. Kh. Samigulin et al. (3) indicated (in 1999), with partial
sequences of the ropC1 gene, that both angiosperms and gymnosperms are
monophyletic and that none of the recent gymnosperms is sister to the
angiosperm.
• S-M Chaw et al. (4) indicate (in 2000), with mitochondrial subunit
rRNA sequences, that gymnosperms are monophyletic; that, in conjecture,
the angiosperms too are monophyletic and substantially older than the
fossil record indicates.
• L.M. Bowe et al. (5) indicated (in 2000), with sequence data of
evolving mitochondrial genes, cox1 and atpA, that extant gymnosperm
genes are monophyletic and that angiosperm origin should be sought
among extinct seed plant groups.
2
|
Introduction
In The Origin of Species, 1859 (6), Charles Darwin wrote:
New Zealand in its endemic
plants is much more closely related to Australia, the nearest mainland,
that to any other region: and this is what might have been expected;
but it is also plainly related to South America, which, although the
next nearest continent, is so enormously remote, that the fact becomes
an anomaly. But this difficulty almost disappears on the view that both
New Zealand, South America and other southern lands were long ago
partially stocked from a nearly intermediate though distant point,
namely from the antarctic islands, when they were clothed with
vegetation, before the commencement of the Glacial period. The
affinity, which, though feeble, I am assured by Dr. [Joseph Dalton]
Hooker is real, between the flora of the south-western corner of
Australia and of the Cape of Good Hope, is a far more remarkable case,
and is at present inexplicable: but this affinity is confined to the
plants, and will, I do not doubt, be some day explained.
On the basis of his 1839-1843 voyages and observations with the
H. M. Discovery ships Erebus and Terror, J. D. Hooker, in 1859 (7),
averred that the evidence of
similar flora on widely separate lands could not be explained without
supposing that these lands once formed a continuous expanse.
Indeed, for Darwin’s “affinity,” Hooker,
perforce, included all biota.
The Grand Puzzle
In what may be termed “one of the grand puzzles of the earth’s
prehistory,” New Zealand and Chile, in common, share native habitat of
earth’s oldest, most primitive flowering plants, side by side with the
earth’s most ancient conifers. New Zealand and Chile are now some 9000
km. apart.
This grand puzzle is resolved herein:
New Zealand was once part of South America on a fully consolidated
Gondwanaland during the late Paleozoic more than 300 million years ago.
A New Zealand block was then adjoined to the southwestern coast of Chile.
From that late Paleozoic date to the present, this essay addresses the
geological and biological relationships of New Zealand to South America
and to Australia.
Premises for the Grand Puzzle
Three premises are explicit to this essay:
• During the late Paleozoic the
southwestern region of South America, as part of Gondwanaland, was
vegetated by a cool temperate rainforest containing
both angiosperms and gymnosperms, both ancestral to the living forests
of present-day Southern Chile.
3
|
• Subsequently, during the
late
Paleozoic, a landmass of obscure contour, here termed the “New Zealand
Plate,” and bearing New Zealand, broke away from South America and
drifted toward the southeastern margin of Australia, while carrying a
cover of indigenous rainforest angiosperms, conifers and other biota.
This landmass came to rest joining margins with Southeastern Australia
about 260 million years ago.
• No natural transfer of
cool-rainforest trees between Australia and New Zealand resulted from
the re-location of New Zealand opposite Australia.
The above premises are buttressed by absence
of fossil evidence of mammals – including Australian indigenous
marsupials – ever endemically inhabiting New Zealand, except for bats.
This thesis posits that except for flying creatures and the effects of
mankind, there has been “total non-connect” of flora and fauna
between Australia and New Zealand; that
New Zealand never was a part of Australia except by submarine attachment;
and that New Zealand is an above-water (outcropping) region of a
volcanic-distorted block, herein termed “New Zealand Plate,” that
rifted off the western coast of South America during the Permo-
Carbonaceous Ice Age, i.e., probably about 280 Ma, and found its
way to the margin of southeastern Australia.
4
|
Part1:
Geological Aspects in Evidence
Cambrian Existence of New
Zealand
There is evidence that New Zealand existed somewhere as
early as the Cambrian era more than 500 million years ago. This
evidence is thus, that the northwest corner of the South Island of New
Zealand strongly indicates that it was once part of a perigondwanic
craton arc system extending landward out to the western coast of South
America. This corner of the South Island is the Kagurangi National
Park, which contains Cambrian and Ordovician material in distributed
belts among Mesozoic bathylithic intrusions. Interestingly, the belts
contain Ordovician fossils such as trilobites and graptolites, among
sedimentary rocks. This evidence is convincing that New Zealand, as
part of a “New Zealand Plate,” is, in fact, of pre-Devonian age.
The New Zealand Plate
Evidence will be cited in support of Devonian Gondwanaland
origin of angiosperms and in support of both angiosperms and conifers
being present over a consolidated Gondwanaland expanse stretching from
Australia to South America prior to 300 Ma. Hence, it is highly
probable that during the late Paleozoic, the New Zealand Plate, along
with contiguous portions of South America, was largely covered by
angiosperm and conifer flora bedded on Cambrian and Ordovician
substrate.
In hypothesis, the New Zealand Plate, a continental block of obscure
conformation, broke away from South America, and drifted off toward
Australia, to be re-configured by geological activity to its present
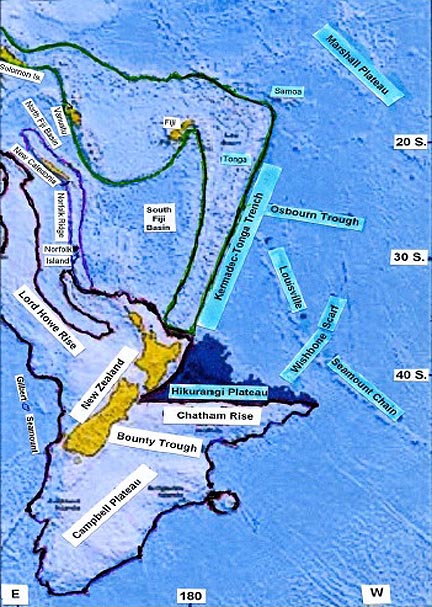
form as shown in Figure 1.
Bathymetric Map of New
Zealand Plate and Surroundings
Figure 1 presents, a bathymetric map of a region of the Southwest
Pacific, in which the present-day New Zealand Plate and New Zealand are
in relationship to their immediate surroundings.
Also shown are New Caledonia, Fiji, Vanuatu, Norfolk Island, and the
Solomon Islands, in context with the Norfolk Ridge, the Lord Howe Rise,
the Gilbert Seamount, and the North and South Fiji Basins. All are
germane
to this presentation. (The basic map is here referenced as K.
Hoernie et al (8). It was obtained in Project Zealandia by scientists
of the Geomar Research Center of Kiel and others, in 2002-2003.)
The New Zealand Plate may have had elongated form when it rifted from
what is now the southwest coast of Chile. While still attached, it
probably reached from 29 S. toward the Fuegan end of Chile. It likely
was the terminus of a South American Cambrian/Ordovician magmatic belt
known as the Rio de la Plata Craton.
Before the rise of the Andes, that craton extended through the
Argentine provinces of San Juan and Mendoza to the coast –and there
into the New Zealand Plate. It is likely that Cambrian and
Ordovician crustal material once covered much surface of New Zealand.
(On a global view, the New Zealand Plate had likely been the extreme
western end of a perigondwanic magmatic Cambrian/Ordovician arc that
included the following cratons: Kaapaal in South Africa;
Kaapvaal-Grunehogna in East Antarctia; Yilgarn in West Australia; and
perhaps even the Lachlan Mountains of New South Wales.
There are also indications that Africa and South America, during
the Cambrian/Ordovician, were united just west of the Kalahari Craton,
an older craton in southern Africa.)
5
|
Figure 1: Bathymetric map showing the present New Zealand Plate
and New Zealand. Also shown are New Caledonia, Fiji, Vanuatu,
tiny Norfolk, and the Solomons; In context the Norfolk Ridge, the
Lord Howe Rise, the Gilbert Seamount, and the North and South
Fiji Basins. All are germane to this presentation. (The basic map was
obtained in Project Zealandia by scientists from the Geomar Research
Center of Kiel and others, in 2002-2003 here referenced to Hoernie et
al.)
 |
Baja South America
In severance from South America, (perceived in Figure 2),
the New Zealand plate took along members of families of ancient,
primitive rainforest trees – leaving behind other members of the same
families. Remarkably, high densities of these relic ancient,
primitive families are alive today at the southern extremity of South
America. This region, which is situated between 37 and 55 S latitude,
is here referred to as “Baja South America,” or simply as “Baja”.
During the late Paleozoic, this region lay south beyond the devastating
ice sheets of the Permo- Carboniferous Ice Age of 300 to 250 million
years ago. Hence, its rainforests were able to survive.
Today these ancient trees come
first into notice on the southern slopes of the Andes from about 37 S
southward. They spread cover over two temperate Chilean rainforests,
the Valdivian, from 41 to 43 S, and the Magellian, from 43 to 55 S. In
these rainforests, 95% of the trees are endemic, and Nothofagus
and Podocarpus are prominent. Baja South America can be
viewed as “a land of living fossil trees,” isolated from other forested
areas.
6
|
Voyage of the New Zealand Plate
In hypothesis, after breaking away from South
America, the
New Zealand Plate drifted past Antarctica and entered into the
Australia domain. The geography of the Gondwanaland continents during
the late Paleozoic suggest a short route, for example, the possible
route, shown in Figure 2 (with B. Windley (9) chart, 1995 as a base)
Today, in order to go from South America past Antarctica to Australia,
a nomad New Zealand would have to travel about 9000 kilometers west
from South America. See Figure 3. During its voyage, and
especially thereafter, the New Zealand Plate has met with conditions of
intense geological foment, attended at times by rapid sea level and
climate changes. Regions have been submerged, obliquely subducted,
compressed, uplifted, transform-faulted, lava-suffused, contorted, or
rifted in geological confrontation with ocean plate tectonics; and
these actions have apparently heightened since the late Cretaceous,
continuing even at present.
|
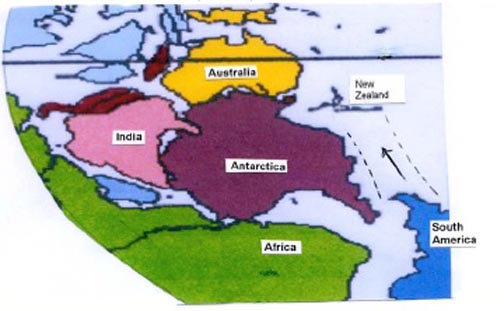 |
Figure 2:
Map illustrating probable oceanic route to Australian margin taken by
New Zealand Plate once broken away from South America. Note that the
journey is shortened considerably by an upside-down South America. The
map is an excerpted sectgion of a paleogeographic map for the start of
the late Paleozoic 410 Ma by Brian Windley in the Evolving Continents, John
Wiley and Sons, Sussex 1995
|
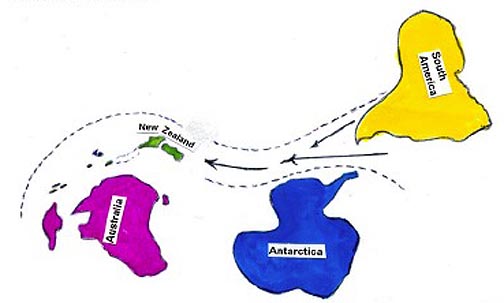 |
Figure 3:
Conceptive view not to scale. Tertiary to present, showing relationship
of New Zealand to Australia. he separation is about 2000 kilometers. To
go past Antarctica from South America today, a nomad New Zealand would
have to drift about 9000 kilometers west.
|
The Above-water Region, New
Zealand
These same geological dynamics applied to above-water New
Zealand, as well. There remains very little of identifiable upper
surface that it brought with it to the vicinity of Australia, (probably
in the late Permian).
The New Zealand Plate may have begun its close
encounter with Australia about 260 million years ago.
7
|
Indications of late-Permian
arrival are offered by volcanic Red Rock near Wellington; by an
ophiolite belt on Dun Mountain near Nelson; and by other rocks here and
there on North and South
Islands.
New Zealand consists of two large islands and a number of smaller
islands that have not drifted away. In area, it is estimated to
represent about 12 % of the entire New Zealand Plate. Its total area is
250,000 sq km, thus indicating a present area of about 2,000,000 sq km
for the New Zealand Plate. Today, the country of New Zealand is about
2100 km from Australia.
The Gilbert Seamount Approach,
New Zealand to Australia
Geological evidence fails to indicate that an overland
passage ever existed between Australia and New Zealand. By evidence
presented below, New Zealand was, at its closest approach, separated
from on-shore Australia by at least 450 km of salt water.
The Gilbert Seamount (in Figure I) is evidence of an undersea terminus
of the continental New Zealand Plate – of a prolongation of the plate
that has imposed a minimum separation of 450 kilometers between the
present New Zealand and Australia, the separation later increasing
accordingly by the interposition of the Tasman Sea (about 80 million
years ago). It implies a formidable rebuttal to any conception of an
overland passage of flora and fauna ever between Australia and New
Zealand.
The Gilbert Seamount is an elevated continental fragment separated from
the adjacent Challenger Plateau. It has an area of about 11,500 sq km.
It is elongated northwest to southeast. It is located at 450 km west of South
Island, New Zealand, and it has been considered continental in origin.
The source of the following regarding the Gilbert Seamount is a paper
by R. A. Wood, 2001, (10).
According to Wood, the Gilbert Seamount appears to be a prolongation of
New Zealand, a western margin, a margin extending westward 450 km
onshore from New Zealand. Both its continuous geological and
morphological connection to the New Zealand mainland support this
assertion, since there is a continuous connection of continental rocks
between the seamount and mainland New Zealand. Hence, here is
indication that, before the intervention of the Tasman Sea about 80
million years ago, New Zealand was in some manner of connection
with the continental margin of Australia at the Gilbert Seamount. Hence, the
New Zealand inland was never closer than 450 kilometers to Australia’s
own southeastern margin.
8
|
To reiterate for its
significance: The above indications clearly argue against
overland passage or land bridge for flora and fauna from Australia to
New Zealand. Contrary to a land connection, they evidence a
450-km underwater shelf, an integral part of the New Zealand
Plate, reaching perhaps all the way to the marginal edge of
Australia from onshore New Zealand and thereby arguing that that
marginal edge of Australia was never closer than 450 km to New
Zealand from the time of the latter’s arrival about 260 million
years ago to the opening of the Tasman Sea.
Geological and Climatological
Circumstances
Concurrent geological and climatological events in context
raise serious objections to the existence of an overland biotic passage
between Australia and New Zealand. One such objection is that sea level
rose worldwide at the beginning of the Cretaceous, from about 130 to
100 Ma. It caused an inland sea in North America from Alaska to the
Gulf of Mexico; and athwart Central Australia, it produced an
epicontinental saltwater incursion that nearly reduced Australia to the
shape of two long islands. The vegetation of Central Australia may have
been totally erased, and the flora of East and West Australia isolated.
Central Australia emerged as a flat desert. On account of its salinity
and aridity, it proved a lasting formidable barrier against the
migrating and intermixing of flora.
A second such objection is that the Tasman Sea opened about 80 Ma, in
the late Cretaceous, and shifted New Zealand and, to a lesser extent,
New Caledonia away from Australia.
A third such objection is that Eastern Australia and Tasmania were
within the Antarctic Circle during almost all of the Cretaceous (until
after the breakaway from Antarctica.) This broad region was then dark
for a part of the year, and had a cold temperate climate. The climate
was, therefore, not conducive to rapid plant diversification and
radiation.
After Australia broke away from Antarctica, the situation of New
Zealand was even more obscure during the Tertiary, due to plate
movement, volcanic activity, and sea level changes. A period of low sea
levels between 53 Ma and 23 Ma may have permitted overland passage
between Norfolk Island and New Caledonia. But New Zealand 29 million
years ago seems to have been flooded, at least partially.
“The information [of the Tertiary] has been so churned up that it will
be difficult to resolve.” Thus, D. H. Tarling, pioneer in the
application of paleomagnetism to continental drift, sums it up in a
personal communication regarding the Southwest Pacific (11).
9
|
Question
of Bathylithic Ascension with Overland Passage
Citing its current position relative to Australia and New
Zealand, it might be suggested that New Caledonia could have been on a
raised conduit or land bridge out from Australia more than 80 million
years ago. There are indications that New Caledonia came into existence
about 100 million years just east of the Lord Howe Rise, the latter
then in marginal attachment with Australia. However, the presence of
New Caledonia gives no answer to the question whether an overland
route, perhaps a sedimentary land bridge, developed, subsequently
permitting access of angiosperms from Australia into New Caledonia, and
from there, perhaps, into New Zealand. There is no evidence of such a
passage – although there is a possibility that such a passage once
existed and could have been subsequently destroyed. Vicariance
describes phenomena of this nature.
The hurdle remains: how to travel overland from New Caledonia to New
Zealand? While New Caledonia is about 600 km closer to Australia than
is New Zealand, today New Caledonia and New Zealand are about 1500 km
apart over the Norfolk Ridge. Again, there may have been overland
passage as far as Norfolk Island. But then, an overland route Norfolk
Island to New Zealand is vastly more difficult to reconcile. These
distances virtually shut the door to previous overland intercourse of
angiosperms.
Ancient Trees on New Caledonia,
Fiji, and Vanuatu
New Caledonia, Fiji, and Vanuatu share the store of
ancient trees with New Zealand and South America – although, in
seeming contradiction of age. New Caledonia is only about 100
million years old; Fiji is less than 80 million; and Vanuatu is less
than 10. Here then, in explication:
Beginning in the late Permian, upon becoming marginally attached to
Australia, the New Zealand Plate entered into a hotbed of geological
activity, mainly volcanism and earthquakes. By the Tertiary, the North
Island of New Zealand had surrendered most of its
Cambrian/Ordovician top cover to volcanism. But some of that extremely
ancient cover had fragmented into numerous large and small island
shards. The opening of the Tasman Sea (about 80 Ma) gave impetus to
these islands drifting away, particularly northward, and finding
allochthonous attachment to plateaus, rises, and seamounts – while
carrying immigrant, prime-condition, ancient and primitive trees of
South American ancestry.
Some fragments found their way north to the narrowed top of the South
Fiji Basin and became parts of Fiji (Figure 1). Other drifting islands
became part of New Caledonia and its offshore islands, entering perhaps
by way of the New Hebrides Trench. Less than 10 million years ago,
other island shards of New Zealand origin participated in forming
Vanuatu by becoming attached to seamounts, by way of the North Fiji
Basin or the New Hebrides Trench.
10
|
Part 2: Biological
Aspects
in Evidence
Introduction
Fossils are by no means the only witnesses to earth
history. The most ancient, most primitive trees and their insect
pollinators – living today– at times are better witnesses where
details of context and continuity of earth history are sought. These
witnesses are especially valuable where fossils are absent. Geography
of living plants and animals interacts herein with tectonics, sea level
change, and climate in accounting for New Zealand’s past and present
relation to Chile.
The most ancient, most primitive trees on earth tell the story of the
separation of the New Zealand Plate from South America. They are
represented here by five families – the angiosperm families,
Nothofagaceae, Proteaceae, and Winteraceae, and the gymnosperm families
Araucariaceae and Podocarpaceae. These families are badges of the unity
of South America and the New Zealand Plate that prevailed more than 300
million years ago. They continue to exist today on continents and
islands now separated by thousands of kilometers. They are described in
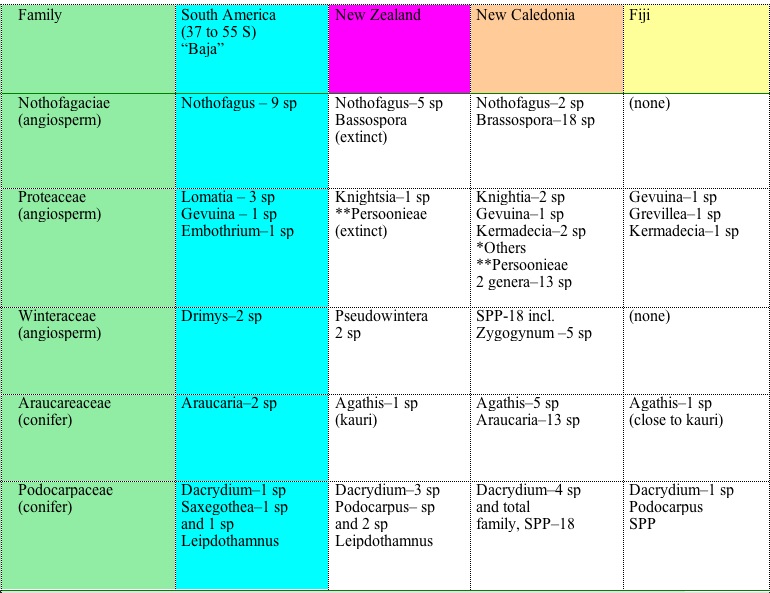
Table 1 below.
Table 1: Description of Five S. Hemisphere Exemplar Plant Families
|
In the late Carboniferous 300 million years ago, Madagascar
was
attached to India and was near to Africa. Moreover, India, Africa,
and Antarctica were either attached or in close proximity to each
other through Australia. To
emphasize, the three
angiosperm and two gymnosperm families cited above more than 300
million years ago are averred here to have occupied the whole of the
southern hemisphere supercontinent Gondwanaland, which was
continuous from South America through Australia.
The above five flora were present during the Carboniferous prior to the
ice invasion, and before 300 million years ago. They occupied a
continuous Gondwanaland expanse stretching from Australia to South
America. Africa had been a main
(middle) section that route of propagation.
11
|
The
Ice Age, the Linchpin of Antiquity
The Permo-Carboniferous Ice Age (also known as the
Gondwanaland Ice Age) began 300 million years ago and lasted 50 million
years. Its magnitude dwarfed recent glaciations like those of the
Pleistocene. According to D.H. Tarling and M.P. Tarling, 1971,
(14), ice fields covered the eastern part of South America above 40 S
latitude and well north into Brazil.
It vastly obliterated the terrestrial life of Africa. In Africa, each
of the above five families was severely affected to this day by the
ice:
1) For the Proteaceae in
Africa, the phylogenetic composition was altered from 14 tribes to
essentially one tribe, the Proteeae.
2) For the Winteraceae in
Africa, all traces were removed.
3) For the Nothofagaceae in
Africa, all traces were removed.
4) For the Araucariaceae in Africa,
all traces were removed.
5) From Africa, the
Podocarpaceae were wiped out; but they managed to make a limited
comeback along the southeastern coast of Africa probably via
Madagascar as the climate later warmed.
Hence, the Permo-Carboniferous Ice Age, begun 300 million years ago, is
a linchpin date for a thesis of Devonian angiosperm and gymnosperm
origin. Both biological and geological evidence have thus been
offered in this essay that the New Zealand Plate is pre-Devonian.
Further biological evidence will be set forth here that the above five
tree families – from the distant past to the present – have
simultaneously occupied New Zealand and Baja South America.
The Five Examplar Tree Families
The five cited tree
families, each deduced to be more than 300 million years old, are today
native to Baja South America and also to New Zealand, New Caledonia,
and Fiji, 9000 kilometers away.
Table 2 summarizes the salient facts:
The Five-Flora Affinities
of Southernmost South America and the Southwest Islands
*Other Proteaceae on New Caledonia
are Eucarpha,
Garnieria, Grevillea, Macadamia, Sleumerodendron, and Stenocarpus.
**The tribe Persoonieae is the most primitive, most ancient of the
Proteaceae. Five of its 8 genera are found in East Australia, evidence
that it was the place of origin of Proteaceae. One genus, Dilobeia, is
found in Madagascar, strongly suggestive of the route of early westward
radiation; and two genera, Beaupreopsis and Beauprea, are found in New
Caledonia. The Persoonieae are deemed here to be the forerunner of the
two subfamilies of Proteaceae.
|
12
|
As a most remarkable
example in Table 2, the same genus,
Gevuina, the
Chile hazel nut tree, of family Proteaceae, is endemic today on
New Caledonia, on Fiji, and also on Vanuatu, – and, amazingly,
on Baja South America, which is about 9000 km and some 300
million years away. Proteaceae cannot tolerate salt water.
The Winteraceae are probably the oldest of the angiosperms. Like the
Proteaceae, they probably originated in Eastern Australia. The
Proteaceae, manifest a “living fossil” genus, Dilobeia, on Madagascar.
As a parallel, the Winteraceae manifest a “living fossil” on
Madagascar. It is the species Takhtajania perrierii. Both of these
ancient living angiosperms mark essentially the same overland route
west from Australia through Indian Madagascar through Africa and into
South America more than 300 million years ago, before the Permo-Carboniferous Ice Age
(300-250 Ma).
In Baja South America today, there are the Winteraceae native
plants, Drimys winteri and Drimys aromatica, 9000 kilometers and 300
million years from New Zealand. |
Table 2 is consistent with an estimate here that the New
Zealand Plate
– bearing the above ancient tree families – broke away from Baja
South America just before or during the Permo-Carboniferous Ice Age.
(The southwestern coast of Chile is noted to have had no ice age
glaciation (14).) It is considered here that the rift may have
occurred at the beginning of the Permian about 280 Ma. The basis
is an early warming and a momentous rise in sea level at coastal
regions of Gondwanaland 280 million years
ago.
In evidence, M.T. Gibbs et al (2002) (15) wrote: “The Permian Period .
. . contains the most recent transformation from a major glaciation to
a generally ice-free state . . . Apparently, the
deglaciation was relatively rapid, being mainly confined to the Early
Permian Sakmarian [285-280 Ma] Stage . . . [A Glossopteris forest cover
replaced ice sheets] and the early and ubiquitous Gandawanan sequence,
from tillites to coal swamp deposits, indicates a major climate
warming . . . ”
The probability is high that at that time (280 Ma) tree-bearing
landmasses other than the New Zealand Plate rifted off elsewhere – for
example, from Antarctica or Australia – ferrying ancient trees to
far-away continental and island sites.
13
|
Part
3: Further Biological and Geological Aspects in Evidence
Introduction
The task ahead is, namely, to provide evidence, example upon example,
in support of Premise 2 that the New Zealand Plate was once attached to
Baja South America (here 37 to 55 south latitude) and in support of
Premise 3 that an overland passage has never existed for flora and
fauna between Australia and New Zealand since the New Zealand Plate
rafted to the vicinity of Australia.
Territorial Family Ties That
Outweigh Time and Distance
The gymnosperm, Lepidothamnus, (family Podocarpaceae in
Table 2) attests in strong support of Premise 2 that three
of its species are very closely related although one is endemic to Baja
South America, and the other two to New Zealand. The following is cited
in 1981 by C. J. Quinn and P. Gadek (16): “Despite the marked geographical
discontinuity, the [three closely related] species are united by their
distinctive cone morphology with its erect ovule, the absence of resin
ducts in the leaves which occur universally elsewhere in the family and
a large number of cupressoid cross-field pits not found elsewhere in
the family. Chemically, these species are also unique in the family,
having cupressuflavone as their major biflavenoid constituent.”
|
Though not mentioned in
Table 2,
impressive angiosperm family ties
belong to the order Laurales and contribute to the ancient rainforest
trees found in both Chile and New Zealand. Of family Monimiaceae, they
are represented by both Laurelia sempervirens and Laureliopsis
philippina, in South America. These species are in indigenous abundance
among the temperate rainforest trees on the Andean slopes and offshore
islands of the upper reaches of Baja South America. They are closely
related to the species Laurelia novae-zelandia present today in New
Zealand.
A remarkable example of affinity of Fiji fauna to South America are two
iguanas. One is the Fiji crested iguana, Brachylophus vitiensis,
indigenous only to Yaduataba Island, near Vanua Levu in Fiji, and to a
few smaller islands. Nearby, the other is the Fiji banded iguana,
Brachylophus fasciatus. Their relatives are opined to live 10,000 km
away in South America and on the Galapagos Islands (once within
reach of South America by archipelagos now under water). No iguanas are
native to Asia, Africa, or Australia.
There are 24 genera of orthocerous weevils endemic to New
Zealand. Relatives of these weevils are found in New Caledonia,
Australia, Sulawesi – and
across the Pacific, in Chile.
On his journey on the Beagle, Darwin described the fungus genus
Cyttaria on Tierra del Fuego and on Tasmania. In New Zealand, three
species of the fungus occur. They are C. gunnii, C. pallida, and C.
nigra. The distribution of the genus follows the distribution of
Nothofagus, which today has 9 species in South America and 5 species in
New Zealand.
Banksia Ericifolia, The Dog that Did Not Bark
A “star witness” to the issue of passage of angiosperms from Australia
to New Zealand is Banksia ericifolia, of family Proteaceae. Banksia
ericifolia offers testimony to Premise 3, in paraphrase: No natural transfer of cool rainforest
trees between Australia and New Zealand resulted from the relocation of
New Zealand opposite Australia. If ever there had been an
overland passage from Southeastern Australia to New Zealand 250 to 80
million years ago, Banksia ericifolia would have had the abundance and
the cold-hardy capability to travel that passage and to flourish in New
Zealand.
Banksia ericifolia, the hearth banksia, bears the imprint of the Primo-
Carboniferous Ice Age in its flowers and in its short, needle-shaped
leaves. It is a survivor of a Permian glacial orogenic foray into New
South Wales 250 million years ago.
14
|
Thus, it is a living example of how the icy cold of 300 to
250 million
years ago altered the angiosperms that were just beyond the reach of
destruction. B. ericifolia came as close to the glaciers of New South
Wales as nature’s chemistry allowed. Ice molded it into an entity hard
and frugal. Its husbandry enabled it to retain only the essentials of
growth and reproduction.
Today B. ericifolia grows in abundance throughout Victoria. It was in high density during the
Cretaceous in Southeast Australia, where its needles were probably a
prime source of nourishment to the cold-adapted members of the dinosaur
infra order Ornithopoda, dominant among the Cretaceous herbivore
dinosaurs of Australia.
Like other Proteaceae, B.ericifolia trees favor coastal hillsides on
the flanks of the ancient Great Dividing Range. Long years ago, they
looked down on the cold water that separated Australia from that advent
New Zealand Plate that, perhaps 260 million years ago, seemed to have
moored offshore. Surely they could have made that polar passage from
Australia to New Zealand, had there been an opportunity. They stood by
to watch as 80 million years ago the widening Tasman Sea took away any
further hope of overland passage.
There is no record of endemic
Banksia ericifolia in New Zealand. Hence, here is a star witness that
an overland passageway never existed from Australia to New Zealand.
Negative evidence as forthright evidence? In “The Hound of the
Baskervilles,” the key evidence was provided by the dog that did not
bark.
Part 4: Mammals,
in
Crucial Support of New Zealand as a
Part of South America
Absence of Mammals
from New Zealand
For mammals, as well as for ancient rainforest trees, Premise 3 of no
overland passage between Australia and New Zealand applies highly
significantly. The beginning of the Carboniferous likely marked the
beginning of the land reptile; and as the period ran its course,
mammal-like reptiles known as therapsids came upon the scene. Later,
the mammalian lineage emerged – roughly 300 million years ago.
At the close of the Carboniferous 280 million years ago, both monotreme
and marsupial mammals roamed Gondwanaland. Eutherian (placental)
mammals may have begun to evolve at that time.
“New Zealand was home to only three land mammals before the arrival of
the [aborigines some 46000 years ago], and all were bats,” according to
T. Flannery and P. Schouten in 2001 (17). And in an earlier discussion
by T. Flannery in 1995 (18): “The New Zealand wattled bat [Chalinolobus
tubercalatus] is the only New Zealand mammal with
Australian affinities . . . ”
Not one
marsupial present there on New Zealand,
or, as it follows, not one on New Caledonia!
15
|
The dates tell the story. There is considerable evidence
that the New
Zealand Plate rifted from South America about 280 million years ago –
about the time that the earliest mammals were just coming into
existence. Hence, there were no mammals other than flying mammals on
New Zealand before the coming of man. Equally significant, from the
time of approach of the New Zealand Plate to Australia about 260
million years, Australian marsupials
never made direct passage to New Zealand, (or indirectly by way
of New Caledonia).
Except for flying creatures and effects of mankind, there has been
“total non-connect” of mammals between Australia and New Zealand.
The above-stated absence of
monotrimes, marsupials and placental mammals from New Zealand is prime
testimony that 1) these animals never crossed over from Australia or
from other parts of Gondwanaland to New Zealand and 2) Australia and
New Zealand were never connected for overland passage.
Part 5: Dinosaur, Bird, and Inter-island Aspects
Were There Dinosaurs in New
Zealand?
The dinosaurs came into existence about 230 million years ago (during
the Triassic). The dinosaurs raise a moot question regarding New
Zealand: Did they ever come to New Zealand? From Australia? Even by way
of New Caledonia about 100 Ma? It is doubtful that there was ever a
native presence of dinosaurs in New Zealand. It is doubtful,
even though dinosaur nesting sites with theropods, ornithopods, and
ankylosaurs undoubtedly existed at Dinosaur Cove in Southeastern
Australia (in Victoria) about 115 million years ago – but at an
extreme sea-travel distance from New Zealand, even for an aquatic
reptile such as a dinosaur.
A few bone fragments of theropods, ornithopods, and ankylosaurs were
found in a stream bed at Hawkes Bay, off North Island, about 1980.
There appears to be no exacting authentication whether these few bone
fragments were brought by nature to that stream bed at a tourist haven;
or, in conjecture, whether there may have been a transplant, perhaps
from Dinosaur Cove. This thesis is not aware of any explicit dinosaur
nesting sites or of any sequence of dinosaur bone discovery and
recording by a professional paleontologist in New Zealand.
16
|
Basic
Distinction among Birds
It would be remiss not to mention that there is at least
one flightless bird that traversed the briny deep from Australia to New
Zealand. It is the penguin – prodigiously-long-distance swimmer. It is
at home today in the coastal waters of New Zealand. There are seven
species on New Zealand shores. Most well-known is Eudyptula minor, the
fairy penguin.
Flying birds evolved from flightless birds of theropod descent. A very
early stage of bird evolution – a stage from feathered theropod to
flightless aquatic bird – goes back about 145 million years ago to the
late Jurassic. Fossil records of 50 million years ago indicate a large
penguin and another large flightless bird, the great auk. True
flightless birds, neither one ever flew. The latter, recently extinct,
was patriarch of the bird family Alcidae, about 21 living
species, all of them illustrating a smooth transition from flightless
to flying bird. Like the penguin, they all are prodigious swimmers.
Attention is given here to the true flightless birds because they are
in sharp distinction to the ratites – the kiwi and the unrelated
extinct moa – natives to New Zealand. Ratites are ground birds that
evolved polyphyletically on ecological opportunity from birds capable
of flight. The ratite amply illustrates a course of evolution in birds,
of different families, that had already evolved flight capability. The trend to flight is reversed in birds
where food is plentiful upon the ground; nesting on the ground is
relatively safe; and there is danger on the wing from predators.
Under such conditions, the body tends to lose its streamlines; wings
become abridged; and flight becomes short and eventually is lost. The
precursors of the kiwi and the moa flew to New Zealand, perhaps from
Australia, perhaps from elsewhere – and found it hospitable.
Inter-island Biotic Commonality
It is relevant to emphasize the close biological relationships among
the islands themselves that stem at least partially from New Zealand.
On New Caledonia, Proteaceae consociate with Nothofagaceae and with the
conifer Araucariaceae. On New Zealand, Nothofagus and Agathis, sister
genus to Araucaria, are together in mixed stands, with Proteaceae
nearby. Both Fiji and New Caledonia have Podocarpus forests.
Two Proteaceae genera are Knightia and Kermandecia. On New Caledonia,
there are two native species of Knightia, the honeysuckle tree; on New
Zealand, there is one. On New Caledonia, there are two native
Kermandecia species; on Fiji and Vanuatu, there is one each. The genera are native nowhere else.
17
|
There is a large bulimoid
snail,
Placostylus, prominent in New Zealand, Norfolk Island, New Caledonia,
Fiji, Vanuatu, and the Solomon Islands, and where land appears on Lord
Howe Rise. Australia and the Louisiades have none. Species are
numerous on Fiji, but Samoa has none. The distribution is said to be in
no way connected with trade winds or ocean currents.
On New Zealand, on both North and South Islands, is found a single
species of an achlorophyllous root saprophyte, an angiosperm of the
family Balanophoraceae. It is Dactylanthus taylori and is found nowhere
else. On New Caledonia is found a single species of Balanophoraceae. It
is Hachettea austrocaledonia and is found nowhere else. The two species
are so closely related anatomically that it has been recommended they
be placed in a separate family of their own, the Dactylanthaceae. As
indicated by their long isolation, they are probably older than other
Balanophoraceae. (Their imputed ancient ancestors, the Amborellaceae,
described below, are now found only on New Caledonia.)
Sole Small-Island Habitations of
Significant Ancient Flora and Fauna
It is startling that among the candidates for the badge of
most ancient angiosperm tree one is solely in New Caledonia and
another, solely in Fiji. One of these is Amborellaceae, a single
species, Amborella tricopoda, found only on New Caledonia. It has been
specifically cited as the most ancient angiosperm and is considered to
be the ancestor of the root holoparasite Balanophoraceae. And
there is found only on Fiji the family Degeneriaceae, two species,
Degeneria vitiensis and Degeneria roseiflora, marked as earliest
primitive by their leaf-like, three-veined stamens and their carpels.
How strange that each among the oldest is found isolated on a separate
small oceanic island! How out of biological and tectonic context it
must seem -the one, only on New Caledonia; the other, only on Fiji!
They could hardly have been imported.
The tuataras belong to the order Rhynchocephalia, a sister to Squamata,
the large, world-wide order of lizards and snakes. Hook-nosed and
lizard-like, the Rhynchocephalia are well-noted in the ancient fossil
record. They are represented today only by the two tuatara species on
tiny islands. They are Sphenodon guntheri and Sphenodon punctatus. They
are native to tiny Lady Alice Island and Stephens Island in New
Zealand. There they find shelter from mammal predators. They are found
nowhere other than in New Zealand. How much out of biological and
tectonic context are the two species of tuataras?
18
|
Conclusions
Three premises of the thesis of late-Paleozoic origin of
the angiosperm and gymnosperm have been presented herein. They are
restated, in paraphrases below, so that the reader may ponder drawing
his or her own conclusions. As Immanuel Kant wrote in 1784, “Have
courage to use your own understanding.”
1. Cool temperate rainforest angiosperm and gymnosperm trees were
conveyed forth on a “New Zealand Plate” rifted from southernmost South
America in the late Paleozoic, probably during the Permo-Carboniferous
Ice Age, leaving close family members behind.
2. Ferried to the vicinity of Australia on the New Zealand Plate, these
ancestral angiosperms and gymnosperms have become today living, though
ancient, tree species of New Zealand and New Caledonia, Fiji, and
Vanuatu.
3. No significant natural transfer of said rainforest trees between
Australia and New Zealand appears to have resulted from the relocation
of the New Zealand Plate offshore of Australia.
The many observations of affinity of ancient biota to both southernmost
South America and the islands of New Zealand, New Caledonia, Fiji, and
Vanuatu are a powerful voice that New Zealand was once attached to
South America.
Highly significantly, the New Zealand absence of Australian marsupials
and placental mammals (excepting bats) proclaims that there never was
an overland passage crossable from Australia to New
Zealand.
L’envoi:
This voyage of Discovery
Ofttimes on perilous
seas
Unfurls an unexpected
sail
In wonderment and beauty.
–Dedicated to J. D. Hooker
19
|
References
1. Troitsky, A.V., Melekhovets, Yu. F.,
Rakhimova,
G.M., Bobrova, V. K., Valiejo-Roman, K.M.,
and Antonov,
A.S., Angiosperm origin . . . deduced from rRNA sequence
comparisons, J.
Mol. Evol.32 (3), 253-61, (1991).
2. Martin, W.F., Lydiate, D., Brinkmann, H., Forkmann, G.,
Saedler,
H., and Rudiger, C.,
Molecular phylogenies in angiosperm
evolution, Mol. Biol. Evol. 10 (1), 140-162 (1993).
3. Samigulin, T. Kh., Martin, W.F., Troitsky, A.V., and Antonov,
A.S.,
Molecular data from the
chloroplast rpoC1 gene suggest a
deep and
distinct dichotomy: gymnosperms (including gnetales)
and angiosperms,
J. Mol.Evol.49 (3), 310-315 (1999).
4. Chaw, S-M., Parkinson, C.L., Cheng, Y., Vincent, T.M., and
Palmer.J.D., Seed plant phylogeny
inferred from all three plant
genomes, PNAS 97, 4086-4091 (2000).
5. Bowe, L.M., Coat, G., and dePamphilis, C.W., Philogeny of seed
plants based on three
genomic compartments, PNAS 97,
4092-4097 (2000).
6. Darwin, C., The Origin of Species: By Means
of Natural Selection, Chapter VIII,
Random House and others.
7. Hooker, J.D., The botany of the Antarctic voyage of H. M.
Discovery
ships Erebus and Terror in
the years 1839-1843, under the command of
Captain Sir James Clark Ross, in Flora Tasmaniae, 3, ~1859.
8. Hoernle, K., Mortimer, N., Werner, R., and Hauff, F., (Eds.)
(2003)
FS Sonne, Fahrtbericht/Cruise
Report SO168 Zealandia “Causes and
Effects of Plume and Rift-related Cretaceous and Cenozoic
Volcanism on
Zealandia,” Geomar Report 133: 127pp+.
9. Windley, B., The Evolving Continent, John Wiley and Sons,
West Sussex, 1995.
10. Wood, R. A., ABLOS
Monaco report, “Finding the Continental Shelf – Examples
from the
New Zealand Region,” ABLOS
01Folder/Wood.
11. Tarling,
D. H., Personal communication to H. Levin, Sept. 28, 2003.
12. Whitmore, T.C., Phytogeography of Eastern Tethys in Wallace’s
Line
and Plate Tectonics, ed.
T.C. Whitmore, Claredon Press,
Oxford,
1981.
13. Whitmore, T.C., Phytogeography of the eastern end of Tethys,
307-311, in Gondwana and
Tethys, eds. M. G. Audley-Charles
and A. Hallam, Geological Society, Oxford Press, 1988.
14. Tarling, D.H. and Tarling, M.P., Continental Drift, a Study
of
the Earth’s Moving Surface,
G. Bell and Sons, London, 1971.
15. Gibbs, M.T., Rees, P.M., Kutzbach, J.E., Ziegler, A.M.,
Behling,
P.J., and Rowley, D.B.,
Simulations of Permian climate and
comparisons with climate-sensitive sediments, The Journal of
Geology,
110, 33-55, (2002).
16. Quinn, C.J. and Gadek, P., Biflavones in Dacrydium sensu
lato, Phytochemistry 20, 677-681 (1981).
17. Flannery, T., and
Schouten, P., A Gap in Nature, p. 167, Atlantic Monthly Press,
2002.
18. Flannery, T., Mammals of the South-West Pacific and Moluccan
Islands, p. 366,
Reed Books, Chatswood, N.S.W
|